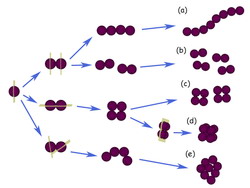
–С–Њ–ї—М—И–µ–є —З–∞—Б—В—М—О, –љ–Њ –љ–µ —Г –≤—Б–µ—Е –≤–Є–і–Њ–≤ (–Є—Б–Ї–ї—О—З–µ–љ–Є–µ —Б–Њ—Б—В–∞–≤–ї—П—О—В –∞–Љ–µ–±—Л, —Н–≤–≥–ї–µ–љ–Њ–≤—Л–µ, –Є–љ—Д—Г–Ј–Њ—А–Є–Є, –і–Є–љ–Њ—Д–ї–∞–≥–µ–ї–ї—П—В—Л, –Љ–љ–Њ–≥–Є–µ –≤–Њ–і–Њ—А–Њ—Б–ї–Є, –љ–µ–Ї–Њ—В–Њ—А—Л–µ –≥—А–Є–±—Л), —П–і–µ—А–љ–∞—П –Њ–±–Њ–ї–Њ—З–Ї–∞ —А–∞–Ј—А—Г—И–∞–µ—В—Б—П –њ—А–Є –Љ–Є—В–Њ–Ј–µ –Є —Б–љ–Њ–≤–∞ –≤–Њ–Ј–љ–Є–Ї–∞–µ—В –њ–Њ—Б–ї–µ –і–µ–ї–µ–љ–Є—П –Ї–ї–µ—В–Њ–Ї. –≠—В–Њ —В–∞–Ї –љ–∞–Ј—Л–≤–∞–µ–Љ—Л–є –Њ—В–Ї—А—Л—В—Л–є —В–Є–њ –Љ–Є—В–Њ–Ј–∞ (—А–Є—Б. 113). –Я—А–Є —Н—В–Њ–Љ –≤ –њ—А–Њ—Д–∞–Ј–µ –њ–Њ –Љ–µ—А–µ –Ї–Њ–љ–і–µ–љ—Б–∞—Ж–Є–Є —Е—А–Њ–Љ–Њ—Б–Њ–Љ —П–і–µ—А–љ–∞—П –Њ–±–Њ–ї–Њ—З–Ї–∞ —В–µ—А—П–µ—В —Б –љ–Є–Љ–Є —Б–≤—П–Ј—М, –∞ –Ј–∞—В–µ–Љ –≤ –љ–µ–є –њ–Њ—П–≤–ї—П—О—В—Б—П —А–∞–Ј—А—Л–≤—Л. –Ю–љ–∞ –њ—А–Є–Њ–±—А–µ—В–∞–µ—В –≤–Є–і –њ–ї–Њ—Б–Ї–Є—Е –Љ–µ–Љ–±—А–∞–љ–љ—Л—Е –≤–∞–Ї—Г–Њ–ї–µ–є, —Ж–Є—Б—В–µ—А–љ. –Т —Н—В–Њ –≤—А–µ–Љ—П —П–і–µ—А–љ—Л–µ –њ–Њ—А—Л –µ—Й–µ –≤–Є–і–љ—Л. –Я–Њ–Ј–і–љ–µ–µ –Њ–љ–Є –Є—Б—З–µ–Ј–∞—О—В. –Т–Њ –≤—А–µ–Љ—П –Љ–Є—В–Њ–Ј–∞ 120 –Љ–Ф–∞ –Ї–Њ–Љ–њ–ї–µ–Ї—Б —П–і–µ—А–љ–Њ–є –њ–Њ—А—Л —А–∞–Ј–±–Є—А–∞–µ—В—Б—П –љ–∞ —Б—Г–±–Ї–Њ–Љ–њ–ї–µ–Ї—Б—Л –њ—А–Є–Љ–µ—А–љ–Њ –њ–Њ 1 –Љ–Ф–∞. –†–∞–Ј–±–Њ—А–Ї–∞ –њ–Њ—А –љ–∞—З–Є–љ–∞–µ—В—Б—П —Б —Д–Њ—Б—Д–Њ—А–Є–ї–Є—А–Њ–≤–∞–љ–Є—П —А—П–і–∞ –љ—Г–Ї–ї–µ–Њ–њ–Њ—А–Є–љ–Њ–≤ –Љ–Є—В–Њ—В–Є—З–µ—Б–Ї–Њ–є cdc2/—Ж–Є–Ї–ї–Є–љ B –Ї–Є–љ–∞–Ј–Њ–є.
–ѓ–і–µ—А–љ–∞—П –Њ–±–Њ–ї–Њ—З–Ї–∞ –њ—А–µ–≤—А–∞—Й–∞–µ—В—Б—П –≤ —Б–Ї–Њ–њ–ї–µ–љ–Є–µ –Љ–µ–ї–Ї–Є—Е –Љ–µ–Љ–±—А–∞–љ–љ—Л—Е –њ—Г–Ј—Л—А—М–Ї–Њ–≤, –Њ–Ї—А—Г–ґ–∞—О—Й–Є—Е –Ј–Њ–љ—Г –±—Л–≤—И–µ–≥–Њ –Є–љ—В–µ—А—Д–∞–Ј–љ–Њ–≥–Њ —П–і—А–∞. –Ґ–∞–Ї–Є–µ –њ—Г–Ј—Л—А—М–Ї–Є –Љ–Њ—А—Д–Њ–ї–Њ–≥–Є—З–µ—Б–Ї–Є –љ–µ–ї—М–Ј—П –Њ—В–ї–Є—З–Є—В—М –Њ—В –і—А—Г–≥–Є—Е –Љ–µ–ї–Ї–Є—Е –≤–∞–Ї—Г–Њ–ї–µ–є –≤ —Ж–Є—В–Њ–њ–ї–∞–Ј–Љ–µ, –Њ–љ–Є –≤–µ—А–Њ—П—В–љ–Њ —Б–ї–Є–≤–∞—О—В—Б—П —Б –≤–∞–Ї—Г–Њ–ї—П–Љ–Є —Н–љ–і–Њ–њ–ї–∞–Ј–Љ–∞—В–Є—З–µ—Б–Ї–Њ–≥–Њ —А–µ—В–Є–Ї—Г–ї—Г–Љ–∞. –Т –Љ–µ—В–∞—Д–∞–Ј–µ –Љ–µ–Љ–±—А–∞–љ–љ—Л–µ —Н–ї–µ–Љ–µ–љ—В—Л —Ж–Є—В–Њ–њ–ї–∞–Ј–Љ—Л –Њ—В—В–µ—Б–љ—П—О—В—Б—П –Ї –њ–µ—А–Є—Д–µ—А–Є—З–µ—Б–Ї–Є–Љ –Ј–Њ–љ–∞–Љ –Ї–ї–µ—В–Њ–Ї –Љ–Є–Ї—А–Њ—В—А—Г–±–Њ—З–Ї–∞–Љ–Є –≤–µ—А–µ—В–µ–љ–∞ –і–µ–ї–µ–љ–Є—П.
–Т –Ї–Њ–љ—Ж–µ –∞–љ–∞—Д–∞–Ј—Л, –Ї–Њ–≥–і–∞ –њ—А–µ–Ї—А–∞—Й–∞–µ—В—Б—П –і–≤–Є–ґ–µ–љ–Є–µ —Е—А–Њ–Љ–Њ—Б–Њ–Љ –Ї –њ—А–Њ—В–Є–≤–Њ–њ–Њ–ї–Њ–ґ–љ—Л–Љ –њ–Њ–ї—О—Б–∞–Љ –Ї–ї–µ—В–Ї–Є, –Љ–µ–Љ–±—А–∞–љ–љ—Л–µ –њ—Г–Ј—Л—А—М–Ї–Є —Ж–Є—В–Њ–њ–ї–∞–Ј–Љ—Л, –Є –≤ –њ–µ—А–≤—Г—О –Њ—З–µ—А–µ–і—М –Љ–µ–Љ–±—А–∞–љ—Л –≥—А–∞–љ—Г–ї—П—А–љ–Њ–≥–Њ —Н–љ–і–Њ–њ–ї–∞–Ј–Љ–∞—В–Є—З–µ—Б–Ї–Њ–≥–Њ —А–µ—В–Є–Ї—Г–ї—Г–Љ–∞ (—Б–Љ. –љ–Є–ґ–µ), –љ–∞—З–Є–љ–∞—О—В –Ї–Њ–љ—В–∞–Ї—В–Є—А–Њ–≤–∞—В—М —Б –њ–Њ–≤–µ—А—Е–љ–Њ—Б—В—М—О —Е—А–Њ–Љ–Њ—Б–Њ–Љ. –≠—В–Є –Ї–Њ–љ—В–∞–Ї—В—Л –њ—А–Њ–Є—Б—Е–Њ–і—П—В —Б–љ–∞—З–∞–ї–∞ –≤ –љ–µ–±–Њ–ї—М—И–Њ–Љ —З–Є—Б–ї–µ —В–Њ—З–µ–Ї, –љ–Њ –Ј–∞—В–µ–Љ –љ–∞—З–Є–љ–∞–µ—В—Б—П –њ–µ—А–µ—Б—В—А–Њ–є–Ї–∞ –Є —А–Њ—Б—В —Н—В–Є—Е –њ–µ—А–≤–Є—З–љ—Л—Е –Ј–∞—З–∞—В–Ї–Њ–≤ —П–і–µ—А–љ–Њ–є –Њ–±–Њ–ї–Њ—З–Ї–Є. –Ю–љ–Є –Є–Ј –Љ–µ–ї–Ї–Є—Е –њ—Г–Ј—Л—А—М–Ї–Њ–≤ –њ—А–µ–≤—А–∞—Й–∞—О—В—Б—П –≤ –њ–ї–Њ—Б–Ї–Є–µ –≤–∞–Ї—Г–Њ–ї–Є, –Ї–Њ—В–Њ—А—Л–µ —А–∞—Б—В—Г—В –≤ —И–Є—А–Є–љ—Г –Є –Њ–±–≤–Њ–ї–∞–Ї–Є–≤–∞—О—В –њ–Њ–≤–µ—А—Е–љ–Њ—Б—В—М –і–µ–Ї–Њ–љ–і–µ–љ—Б–Є—А—Г—О—Й–Є—Е—Б—П —Е—А–Њ–Љ–Њ—Б–Њ–Љ. –£—З–∞—Б—В–Ї–Є —В–∞–Ї–Є—Е —А–∞—Б—В—Г—Й–Є—Е –њ–ї–Њ—Б–Ї–Є—Е –Љ–µ–Љ–±—А–∞–љ–љ—Л—Е –Љ–µ—И–Ї–Њ–≤ —Б–ї–Є–≤–∞—О—В—Б—П, –Ј–∞–Љ—Л–Ї–∞—П –Є –Њ—В–≥–Њ—А–∞–ґ–Є–≤–∞—П —Б–Њ–і–µ—А–ґ–Є–Љ–Њ–µ –љ–Њ–≤–Њ–≥–Њ –Є–љ—В–µ—А—Д–∞–Ј–љ–Њ–≥–Њ —П–і—А–∞. –Ш–љ—В–µ—А–µ—Б–љ–Њ, —З—В–Њ —П–і–µ—А–љ—Л–µ –њ–Њ—А—Л –њ–Њ—П–≤–ї—П—О—В—Б—П –љ–∞ —Б–∞–Љ—Л—Е —А–∞–љ–љ–Є—Е —Н—В–∞–њ–∞—Е —А–µ–Ї–Њ–љ—Б—В—А—Г–Ї—Ж–Є–Є —П–і–µ—А–љ–Њ–є –Њ–±–Њ–ї–Њ—З–Ї–Є, –Ї–Њ–≥–і–∞ –і–≤–Њ–є–љ—Л–µ –Љ–µ–Љ–±—А–∞–љ–љ—Л–µ —Ж–Є—Б—В–µ—А–љ—Л –µ—Й–µ –љ–µ —Б–Њ–Љ–Ї–љ—Г–ї–Є—Б—М –Є —Д–∞–Ї—В–Є—З–µ—Б–Ї–Є –љ–Є—З–µ–≥–Њ –љ–µ —А–∞–Ј–і–µ–ї—П—О—В.
–Я—А–Є —А–µ–Ї–Њ–љ—Б—В—А—Г–Ї—Ж–Є–Є —П–і–µ—А–љ–Њ–є –Њ–±–Њ–ї–Њ—З–Ї–Є –њ—А–Њ–Є—Б—Е–Њ–і–Є—В —Б–±–Њ—А–Ї–∞ —П–і–µ—А–љ—Л—Е –њ–Њ—А. –Ю–љ–∞ –љ–∞—З–Є–љ–∞–µ—В—Б—П —Б –Њ–±—А–∞–Ј–Њ–≤–∞–љ–Є—П —П–Љ–Ї–Є –њ—А–Є —Б–ї–Є—П–љ–Є–Є –≤–љ–µ—И–љ–µ–є –Є –≤–љ—Г—В—А–µ–љ–љ–µ–є —П–і–µ—А–љ–Њ–є –Љ–µ–Љ–±—А–∞–љ—Л, –Ї–Њ—В–Њ—А–∞—П –Ј–∞—В–µ–Љ –њ—А–µ–≤—А–∞—Й–∞–µ—В—Б—П –≤ –Њ—В–≤–µ—А—Б—В–Є–µ. –Т —Н—В–Њ–Љ –њ—А–Њ—Ж–µ—Б—Б–µ –њ—А–Є–љ–Є–Љ–∞—О—В —Г—З–∞—Б—В–Є–µ –Є–љ—В–µ–≥—А–∞–ї—М–љ—Л–µ –±–µ–ї–Ї–Є gp 210 –Є POM 121, –Ї–Њ—В–Њ—А—Л–µ –≤–њ–Њ—Б–ї–µ–і—Б—В–≤–Є–Є –±—Г–і—Г—В –Ј–∞–Ї—А–µ–њ–ї—П—В—М –ѓ–Я–Ъ –љ–∞ –Љ–µ–Љ–±—А–∞–љ–∞—Е.
–Ч–∞ —Н—В–Є–Љ —Б–ї–µ–і—Г–µ—В –њ–Њ—П–≤–ї–µ–љ–Є–µ –≤–љ—Г—В—А–µ–љ–љ–Є—Е —Б—В—А—Г–Ї—В—Г—А –ѓ–Я–Ъ: –Ї–Њ–Љ–њ–ї–µ–Ї—Б –Ї–Њ–ї—М—Ж–∞, —Б–њ–Є—Ж, –і–Њ–±–∞–≤–ї–µ–љ–Є–µ –Ј–≤–µ–Ј–і—З–∞—В–Њ–≥–Њ –Ї–Њ–ї—М—Ж–∞ –Є –і—А—Г–≥–Є—Е —Б—В—А—Г–Ї—В—Г—А, –Є, –љ–∞–Ї–Њ–љ–µ—Ж, —Д–Є–ї–∞–Љ–µ–љ—В–Њ–≤.
–£ –љ–µ–Ї–Њ—В–Њ—А—Л—Е –љ–Є–Ј—И–Є—Е –Њ—А–≥–∞–љ–Є–Ј–Љ–Њ–≤ –≤ —Б–ї—Г—З–∞–µ –Ј–∞–Ї—А—Л—В–Њ–≥–Њ –Љ–Є—В–Њ–Ј–∞ —П–і–µ—А–љ–∞—П –Њ–±–Њ–ї–Њ—З–Ї–∞ –љ–µ –Є—Б—З–µ–Ј–∞–µ—В, –Њ–љ–∞ –≤ –Ј–Њ–љ–µ —П–і–µ—А–љ–Њ–є –њ–µ—А–µ—В—П–ґ–Ї–Є –Ј–∞–Љ—Л–Ї–∞–µ—В—Б—П, —З—В–Њ –њ—А–Є–≤–Њ–і–Є—В –Ї –Њ–±—А–∞–Ј–Њ–≤–∞–љ–Є—О –і–≤—Г—Е –љ–Њ–≤—Л—Е —П–і–µ—А. –Ч–і–µ—Б—М —Г—З–∞—Б—В–Є–µ —П–і–µ—А–љ–Њ–є –Њ–±–Њ–ї–Њ—З–Ї–Є –≤ –і–µ–ї–µ–љ–Є–Є –Ї–ї–µ—В–Ї–Є –Ј–∞–Ї–ї—О—З–∞–µ—В—Б—П –≤ —В–Њ–Љ, —З—В–Њ –љ–∞ –љ–µ–є –Ј–∞–Ї—А–µ–њ–ї–µ–љ—Л —Е—А–Њ–Љ–Њ—Б–Њ–Љ—Л, –Є –Њ–љ–∞, –њ–Њ-–≤–Є–і–Є–Љ–Њ–Љ—Г, –њ—А–Є–љ–Є–Љ–∞–µ—В —Г—З–∞—Б—В–Є–µ –≤ –Є–љ–і—Г–Ї—Ж–Є–Є –Њ–±—А–∞–Ј–Њ–≤–∞–љ–Є—П –Љ–Є–Ї—А–Њ—В—А—Г–±–Њ—З–µ–Ї, –љ–µ–Њ–±—Е–Њ–і–Є–Љ—Л—Е –њ—А–Є –і–µ–ї–µ–љ–Є–Є –Ї–ї–µ—В–Њ–Ї.
–Я–Њ-–≤–Є–і–Є–Љ–Њ–Љ—Г, –і–ї—П —А–µ–Ї–Њ–љ—Б—В—А—Г–Ї—Ж–Є–Є —П–і–µ—А–љ–Њ–є –Њ–±–Њ–ї–Њ—З–Ї–Є –љ–µ–Њ–±—Е–Њ–і–Є–Љ—Л–Љ —Г—Б–ї–Њ–≤–Є–µ–Љ —П–≤–ї—П–µ—В—Б—П –і–µ–Ї–Њ–љ–і–µ–љ—Б–∞—Ж–Є—П —Е—А–Њ–Љ–Њ—Б–Њ–Љ. –С—Л–ї–Њ –њ–Њ–Ї–∞–Ј–∞–љ–Њ, —З—В–Њ –µ—Б–ї–Є –≤—Л–Ј–≤–∞—В—М –њ—А–µ–ґ–і–µ–≤—А–µ–Љ–µ–љ–љ—Г—О –і–µ–Ї–Њ–љ–і–µ–љ—Б–∞—Ж–Є—О –Љ–µ—В–∞—Д–∞–Ј–љ—Л—Е —Е—А–Њ–Љ–Њ—Б–Њ–Љ, —В–Њ –Њ–љ–Є –Њ—З–µ–љ—М –±—Л—Б—В—А–Њ –Ї–Њ–љ—В–∞–Ї—В–Є—А—Г—О—В —Б –Љ–µ–Љ–±—А–∞–љ–љ—Л–Љ–Є –њ—Г–Ј—Л—А—М–Ї–∞–Љ–Є –Є –Њ–і–µ–≤–∞—О—В—Б—П –Ї–∞–ґ–і–∞—П —Б–≤–Њ–µ–є –Њ—В–і–µ–ї—М–љ–Њ —П–і–µ—А–љ–Њ–є –Њ–±–Њ–ї–Њ—З–Ї–Њ–є, –≤—Б–ї–µ–і—Б—В–≤–Є–µ —З–µ–≥–Њ –≤ –Ї–ї–µ—В–Ї–µ –≤–Њ–Ј–љ–Є–Ї–∞–µ—В –Љ–љ–Њ–ґ–µ—Б—В–≤–Њ —В–∞–Ї –љ–∞–Ј—Л–≤–∞–µ–Љ—Л—Е –Љ–Є–Ї—А–Њ—П–і–µ—А, –Ї–∞–ґ–і–Њ–µ –Є—Е –Ї–Њ—В–Њ—А—Л—Е –≤–Њ–Ј–љ–Є–Ї–ї–Њ –Є–Ј –Њ–і–љ–Њ–є —Е—А–Њ–Љ–Њ—Б–Њ–Љ—Л.
–° –і—А—Г–≥–Њ–є —Б—В–Њ—А–Њ–љ—Л, –Љ–Њ–ґ–љ–Њ —Н–Ї—Б–њ–µ—А–Є–Љ–µ–љ—В–∞–ї—М–љ–Њ –≤—Л–Ј–≤–∞—В—М —А–∞–Ј–±–Њ—А–Ї—Г —П–і–µ—А–љ–Њ–є –Њ–±–Њ–ї–Њ—З–Ї–Є —Г –Є–љ—В–µ—А—Д–∞–Ј–љ–Њ–≥–Њ —П–і—А–∞. –≠—В–Њ –њ—А–Њ–Є—Б—Е–Њ–і–Є—В, –µ—Б–ї–Є —Б–ї–Є—В—М –≤ –Ї—Г–ї—М—В—Г—А–µ —В–Ї–∞–љ–Є –і–≤–µ –Ї–ї–µ—В–Ї–Є –љ–∞ —А–∞–Ј–љ—Л—Е —Б—В–∞–і–Є—П—Е –Ї–ї–µ—В–Њ—З–љ–Њ–≥–Њ —Ж–Є–Ї–ї–∞ –Є –њ–Њ–ї—Г—З–Є—В—М —В.–љ. –≥–µ—В–µ—А–Њ–Ї–∞—А–Є–Њ–љ, –≥–і–µ –Њ–і–љ–Њ –Є–Ј —П–і–µ—А –±—Г–і–µ—В –љ–∞—Е–Њ–і–Є—В—М—Б—П –≤ –Є–љ—В–µ—А—Д–∞–Ј–µ, –∞ –і—А—Г–≥–Њ–µ –±—Л—В—М –≤ –≤–Є–і–µ –Љ–Є—В–Њ—В–Є—З–µ—Б–Ї–Є—Е —Е—А–Њ–Љ–Њ—Б–Њ–Љ –≤ –Љ–µ—В–∞—Д–∞–Ј–µ. –Т —Н—В–Њ–Љ —Б–ї—Г—З–∞–µ –≤ –Є–љ—В–µ—А—Д–∞–Ј–љ–Њ–Љ —П–і—А–µ –љ–∞—З–Є–љ–∞–µ—В –Ї–Њ–љ–і–µ–љ—Б–Є—А–Њ–≤–∞—В—М—Б—П —Е—А–Њ–Љ–∞—В–Є–љ, –Њ–±—А–∞–Ј—Г—О—В—Б—П –њ—А–µ–ґ–і–µ–≤—А–µ–Љ–µ–љ–љ–Њ –Ї–Њ–љ–і–µ–љ—Б–Є—А–Њ–≤–∞–љ–љ—Л–µ —Е—А–Њ–Љ–Њ—Б–Њ–Љ—Л, –∞ —П–і–µ—А–љ–∞—П –Њ–±–Њ–ї–Њ—З–Ї–∞ –Є—Б—З–µ–Ј–љ–µ—В —В–∞–Ї –ґ–µ –Ї–∞–Ї –≤–Њ –≤—А–µ–Љ—П –љ–Њ—А–Љ–∞–ї—М–љ–Њ–≥–Њ –Љ–Є—В–Њ–Ј–∞ (—А–Є—Б. 114). –≠—В–Є –і–∞–љ–љ—Л–µ –≥–Њ–≤–Њ—А—П—В –Њ —В–Њ–Љ, —З—В–Њ –≤ —Ж–Є—В–Њ–њ–ї–∞–Ј–Љ–µ –Љ–Є—В–Њ—В–Є—З–µ—Б–Ї–Њ–є –Ї–ї–µ—В–Ї–Є —Б—Г—Й–µ—Б—В–≤—Г—О—В –Ї–∞–Ї–Є–µ-—В–Њ —Д–∞–Ї—В–Њ—А—Л, –≤—Л–Ј—Л–≤–∞—О—Й–Є–µ –Ї–∞–Ї –Ї–Њ–љ–і–µ–љ—Б–∞—Ж–Є—О —Е—А–Њ–Љ–Њ—Б–Њ–Љ, —В–∞–Ї –Є –њ–∞—А–∞–ї–ї–µ–ї—М–љ—Л–є —Н—В–Њ–Љ—Г –њ—А–Њ—Ж–µ—Б—Б —А–∞—Б–њ–∞–і–∞ —П–і–µ—А–љ–Њ–є –Њ–±–Њ–ї–Њ—З–Ї–Є.
–°—Е–Њ–і–љ–∞—П –і–Є–љ–∞–Љ–Є–Ї–∞ —Б–Њ–≤–њ–∞–і–µ–љ–Є—П –њ—А–Њ—Ж–µ—Б—Б–Њ–≤ –њ–µ—А–µ—Б—В—А–Њ–є–Ї–Є —Е—А–Њ–Љ–Њ—Б–Њ–Љ –Є —П–і–µ—А–љ–Њ–є –Њ–±–Њ–ї–Њ—З–Ї–Є –љ–∞–±–ї—О–і–∞–µ—В—Б—П –Є –≤ –і—А—Г–≥–Њ–є —Б–Є—Б—В–µ–Љ–µ, –≤ —Ж–Є—В–Њ–њ–ї–∞–Ј–Љ–µ –Њ–Њ—Ж–Є—В–Њ–≤ –Є–ї–Є –≤ –±–µ—Б–Ї–ї–µ—В–Њ—З–љ—Л—Е —Ж–Є—В–Њ–њ–ї–∞–Ј–Љ–∞—В–Є—З–µ—Б–Ї–Є—Е —Н–Ї—Б—В—А–∞–Ї—В–∞—Е –Њ–Њ—Ж–Є—В–Њ–≤. –Ґ–∞–Ї –µ—Б–ї–Є –≤ —Ж–Є—В–Њ–њ–ї–∞–Ј–Љ—Г –Њ–Њ—Ж–Є—В–∞ –∞–Љ—Д–Є–±–Є–є –љ–∞ —Б—В–∞–і–Є–Є –Љ–µ—В–∞—Д–∞–Ј—Л –Є–љ—К–µ—Ж–Є—А–Њ–≤–∞—В—М –≤—Л–і–µ–ї–µ–љ–љ—Л–µ –Є–љ—В–µ—А—Д–∞–Ј–љ—Л–µ —П–і—А–∞, —В–Њ –Є—Е —П–і–µ—А–љ–∞—П –Њ–±–Њ–ї–Њ—З–Ї–∞ —А–∞–Ј–±–Є—А–∞–µ—В—Б—П, –∞ —Е—А–Њ–Љ–∞—В–Є–љ –Ї–Њ–љ–і–µ–љ—Б–Є—А—Г–µ—В—Б—П –≤ –≤–Є–і–µ –Љ–Є—В–Њ—В–Є—З–µ—Б–Ї–Є—Е —Е—А–Њ–Љ–Њ—Б–Њ–Љ. –Х—Б–ї–Є –ґ–µ –≤ –Њ–Њ—Ж–Є—В –љ–∞ —Б—В–∞–і–Є–Є –Є–љ—В–µ—А—Д–∞–Ј—Л –≤–≤–µ—Б—В–Є –Љ–Є—В–Њ—В–Є—З–µ—Б–Ї–Є–µ —Е—А–Њ–Љ–Њ—Б–Њ–Љ—Л, —В–Њ –Њ–љ–Є –љ–∞—З–Є–љ–∞—О—В –і–µ–Ї–Њ–љ–і–µ–љ—Б–Є—А–Њ–≤–∞—В—М—Б—П, –њ–Њ—П–≤–ї—П—О—В—Б—П –Љ–љ–Њ–ґ–µ—Б—В–≤–µ–љ–љ—Л–µ –Љ–µ–ї–Ї–Є–µ –≤–∞–Ї—Г–Њ–ї–Є, –Ї–Њ—В–Њ—А—Л–µ —Б–ї–Є–≤–∞—П—Б—М –і—А—Г–≥ —Б –і—А—Г–≥–Њ–Љ, –Њ–±—А–∞–Ј—Г—О—В —П–і–µ—А–љ—Л–µ –Њ–±–Њ–ї–Њ—З–Ї–Є. –Ш–љ—В–µ—А–µ—Б–љ–Њ, —З—В–Њ –≤ —Ж–Є—В–Њ–њ–ї–∞–Ј–Љ—Г –Є–љ—В–µ—А—Д–∞–Ј–љ–Њ–≥–Њ –Њ–Њ—Ж–Є—В–∞ –Љ–Њ–ґ–љ–Њ –≤–≤–µ—Б—В–Є –і–∞–ґ–µ —З—Г–ґ–µ—А–Њ–і–љ—Г—О —З–Є—Б—В—Г—О –Ф–Э–Ъ, –Ї–Њ—В–Њ—А–∞—П, —Б–≤—П–Ј—Л–≤–∞—П—Б—М —Б –≥–Є—Б—В–Њ–љ–∞–Љ–Є –≤ —Ж–Є—В–Њ–њ–ї–∞–Ј–Љ–µ, –Њ–±—А–∞–Ј—Г–µ—В —Е—А–Њ–Љ–∞—В–Є–љ–Њ–≤—Л–µ –≥–ї—Л–±–Ї–Є, –Ї–Њ—В–Њ—А—Л–µ –≤ —Б–≤–Њ—О –Њ—З–µ—А–µ–і—М –Њ–і–µ–≤–∞—О—В—Б—П —П–і–µ—А–љ—Л–Љ–Є –Њ–±–Њ–ї–Њ—З–Ї–∞–Љ–Є –Є –њ—А–µ–≤—А–∞—Й–∞—О—В—Б—П –≤ –Љ–Є–Ї—А–Њ—П–і—А–∞.
–≠—В–Є —Н–Ї—Б–њ–µ—А–Є–Љ–µ–љ—В–∞–ї—М–љ—Л–µ –њ—А–Є–µ–Љ—Л –≤–Љ–µ—Б—В–µ —Б –Љ–µ—В–Њ–і–Њ–Љ –Є–Љ–Љ—Г–љ–Њ—Д–ї—Г–Њ—А–µ—Б—Ж–µ–љ—Ж–Є–Є –њ–Њ–Ј–≤–Њ–ї–Є–ї–Є –њ—А–Њ—Б–ї–µ–і–Є—В—М —Б—Г–і—М–±—Г –Љ–љ–Њ–≥–Є—Е –±–µ–ї–Ї–Њ–≤ —П–і–µ—А–љ–Њ–є –Њ–±–Њ–ї–Њ—З–Ї–Є –≤–Њ –≤—А–µ–Љ—П –Љ–Є—В–Њ–Ј–∞. –Я–Њ–і—А–Њ–±–љ–Њ –Є–Ј—Г—З–µ–љ–∞ —Б—Г–і—М–±–∞ –ї–∞–Љ–Є–љ–Њ–≤. –С—Л–ї–Њ –љ–∞–є–і–µ–љ–Њ, —З—В–Њ —Д–Є–±—А–Њ–Ј–љ—Л–є —Б–ї–Њ–є –ї–∞–Љ–Є–љ–Њ–≤ –і–µ–њ–Њ–ї–Є–Љ–µ—А–Є–Ј—Г–µ—В—Б—П –њ–∞—А–∞–ї–ї–µ–ї—М–љ–Њ —А–∞—Б–њ–∞–і—Г —П–і–µ—А–љ—Л—Е –Љ–µ–Љ–±—А–∞–љ –Є –Ї–Њ–љ–і–µ–љ—Б–∞—Ж–Є–Є —Е—А–Њ–Љ–∞—В–Є–љ–∞. –≠—В–Њ–Љ—Г –њ—А–µ–і—И–µ—Б—В–≤—Г–µ—В –Њ–±–Є–ї—М–љ–Њ–µ (–≤ 7 —А–∞–Ј –≤—Л—И–µ, —З–µ–Љ –≤ –Є–љ—В–µ—А—Д–∞–Ј–µ) —Д–Њ—Б—Д–Њ—А–Є–ї–Є—А–Њ–≤–∞–љ–Є–µ –ї–∞–Љ–Є–љ–Њ–≤. –Ы–∞–Љ–Є–љ—Л A –Є C –њ—А–Є —Н—В–Њ–Љ –і–µ–њ–Њ–ї–Є–Љ–µ—А–Є–Ј—Г—О—В—Б—П –і–Њ –і–Є–Љ–µ—А–Њ–≤ –Є —В–µ—В—А–∞–Љ–µ—А–Њ–≤ –Є, –њ–µ—А–µ—Е–Њ–і—П –≤ —А–∞—Б—В–≤–Њ—А–Є–Љ–Њ–µ —Б–Њ—Б—В–Њ—П–љ–Є–µ, —А–∞–≤–љ–Њ–Љ–µ—А–љ–Њ —А–∞—Б–њ—А–µ–і–µ–ї—П—О—В—Б—П –≤ —Ж–Є—В–Њ–њ–ї–∞–Ј–Љ–µ –≤–љ–µ —Б–≤—П–Ј–Є —Б –і—А—Г–≥–Є–Љ–Є —Б—В—А—Г–Ї—В—Г—А–∞–Љ–Є. –Ы–∞–Љ–Є–љ B —В–Њ–ґ–µ –і–µ–њ–Њ–ї–Є–Љ–µ—А–Є–Ј—Г–µ—В—Б—П –і–Њ –Њ–ї–Є–≥–Њ–Љ–µ—А–Њ–≤, –љ–Њ –Њ—Б—В–∞–µ—В—Б—П —Б–≤—П–Ј–∞–љ–љ—Л–Љ —Б –Љ–µ–Љ–±—А–∞–љ–љ—Л–Љ–Є –њ—Г–Ј—Л—А—М–Ї–∞–Љ–Є, –≤–Њ–Ј–љ–Є–Ї—И–Є–Љ–Є –Є–Ј —П–і–µ—А–љ–Њ–є –Њ–±–Њ–ї–Њ—З–Ї–Є.
–Я—А–Є —Б–±–Њ—А–Ї–µ —П–і–µ—А–љ–Њ–є –Њ–±–Њ–ї–Њ—З–Ї–Є –≤ —В–µ–ї–Њ—Д–∞–Ј–µ –±–µ–ї–Ї–Є –ї–∞–Љ–Є–љ—Л –Є–Љ–Љ—Г–љ–Њ—Е–Є–Љ–Є—З–µ—Б–Ї–Є –љ–∞—З–Є–љ–∞—О—В –≤—Л—П–≤–ї—П—В—М—Б—П –≤ —Ж–µ–љ—В—А–Њ–Љ–µ—А–љ—Л—Е –Є —В–µ–ї–Њ–Љ–µ—А–љ—Л—Е —Г—З–∞—Б—В–Ї–∞—Е —Е—А–Њ–Љ–Њ—Б–Њ–Љ, —В–∞–Љ –ґ–µ –Њ–±–љ–∞—А—Г–ґ–Є–≤–∞—О—В—Б—П –њ–µ—А–≤—Л–µ –њ—А–Є–Ј–љ–∞–Ї–Є –Њ–±—А–∞–Ј–Њ–≤–∞–љ–Є—П –љ–Њ–≤–Њ–є —П–і–µ—А–љ–Њ–є –Њ–±–Њ–ї–Њ—З–Ї–Є. –Ґ–∞–Љ –ґ–µ –љ–∞–Ї–∞–њ–ї–Є–≤–∞—О—В—Б—П –∞–љ—В–Є—В–µ–ї–∞ –Ї –±–µ–ї–Ї–∞–Љ –њ–Њ—А–Њ–≤–Њ–≥–Њ –Ї–Њ–Љ–њ–ї–µ–Ї—Б–∞. –Т –±–µ—Б–Ї–ї–µ—В–Њ—З–љ–Њ–є —Б–Є—Б—В–µ–Љ–µ —Ж–Є—В–Њ–њ–ї–∞–Ј–Љ–∞—В–Є—З–µ—Б–Ї–Њ–≥–Њ —Н–Ї—Б—В—А–∞–Ї—В–∞ –Њ–Њ—Ж–Є—В–Њ–≤ –±—Л–ї–Њ –њ–Њ–Ї–∞–Ј–∞–љ–Њ, —З—В–Њ –∞—Б—Б–Њ—Ж–Є–∞—Ж–Є—П —А–∞—Б—В–≤–Њ—А–Є–Љ—Л—Е –≤ –Љ–Є—В–Њ–Ј–µ –ї–∞–Љ–Є–љ–Њ–≤ A –Є C –њ—А–Њ–Є—Б—Е–Њ–і–Є—В –љ–µ–Ј–∞–≤–Є—Б–Є–Љ–Њ –Њ—В –ї–∞–Љ–Є–љ–∞ B. –Ю–Ї–∞–Ј–∞–ї–Њ—Б—М, —З—В–Њ –µ—Б–ї–Є —Б–Є—Б—В–µ–Љ—Г —А–µ–Ї–Њ–љ—Б—В—А—Г–Ї—Ж–Є–Є —П–і–µ—А–љ–Њ–є –Њ–±–Њ–ї–Њ—З–Ї–Є –ї–Є—И–Є—В—М –ї–∞–Љ–Є–љ–∞ B, —В–Њ –ї–∞–Љ–Є–љ—Л A –Є C —Б–≤—П–Ј—Л–≤–∞—О—В—Б—П —Б –њ–Њ–≤–µ—А—Е–љ–Њ—Б—В—М—О —Е—А–Њ–Љ–Њ—Б–Њ–Љ, –љ–Њ —Б–±–Њ—А–Ї–Є —П–і–µ—А–љ–Њ–є –Њ–±–Њ–ї–Њ—З–Ї–Є –љ–µ –њ—А–Њ–Є—Б—Е–Њ–і–Є—В. –Т —Н–Ї—Б—В—А–∞–Ї—В–µ, –ї–Є—И–µ–љ–љ–Њ–Љ –ї–∞–Љ–Є–љ–Њ–≤ A –Є C, –ї–∞–Љ–Є–љ B —Б–≤—П–Ј—Л–≤–∞–µ—В—Б—П —Б —Е—А–Њ–Љ–Њ—Б–Њ–Љ–∞–Љ–Є, –љ–Њ –љ–Њ—А–Љ–∞–ї—М–љ–∞—П —П–і–µ—А–љ–∞—П –Њ–±–Њ–ї–Њ—З–Ї–∞ —В–∞–Ї –ґ–µ –љ–µ —Д–Њ—А–Љ–Є—А—Г–µ—В—Б—П.